
Another interesting type of fluorescent proteins are Fluorescent Timers which are able to change their emission wavelength over time. The following article will give an overview of a selection of these extraordinary fluorescent proteins.

Photoactivatable proteins
Photoactivatable proteins can be "switched on" from a low fluorescent state to a higher fluorescent state. This switch is happening during less than a second by applying a short light pulse in the violet/blue spectrum and allows detection of dynamic cellular processes. Directed photoactivation of molecules in a region of interest (ROI) can be used to monitor the movement of these activated proteins inside the cell. Whereas in other pulse-chase experimental setups like FRAP one cannot distinguish between proteins which are re-entering the ROI and newly synthesized molecules, photoactivation is one way to bypass this problem.
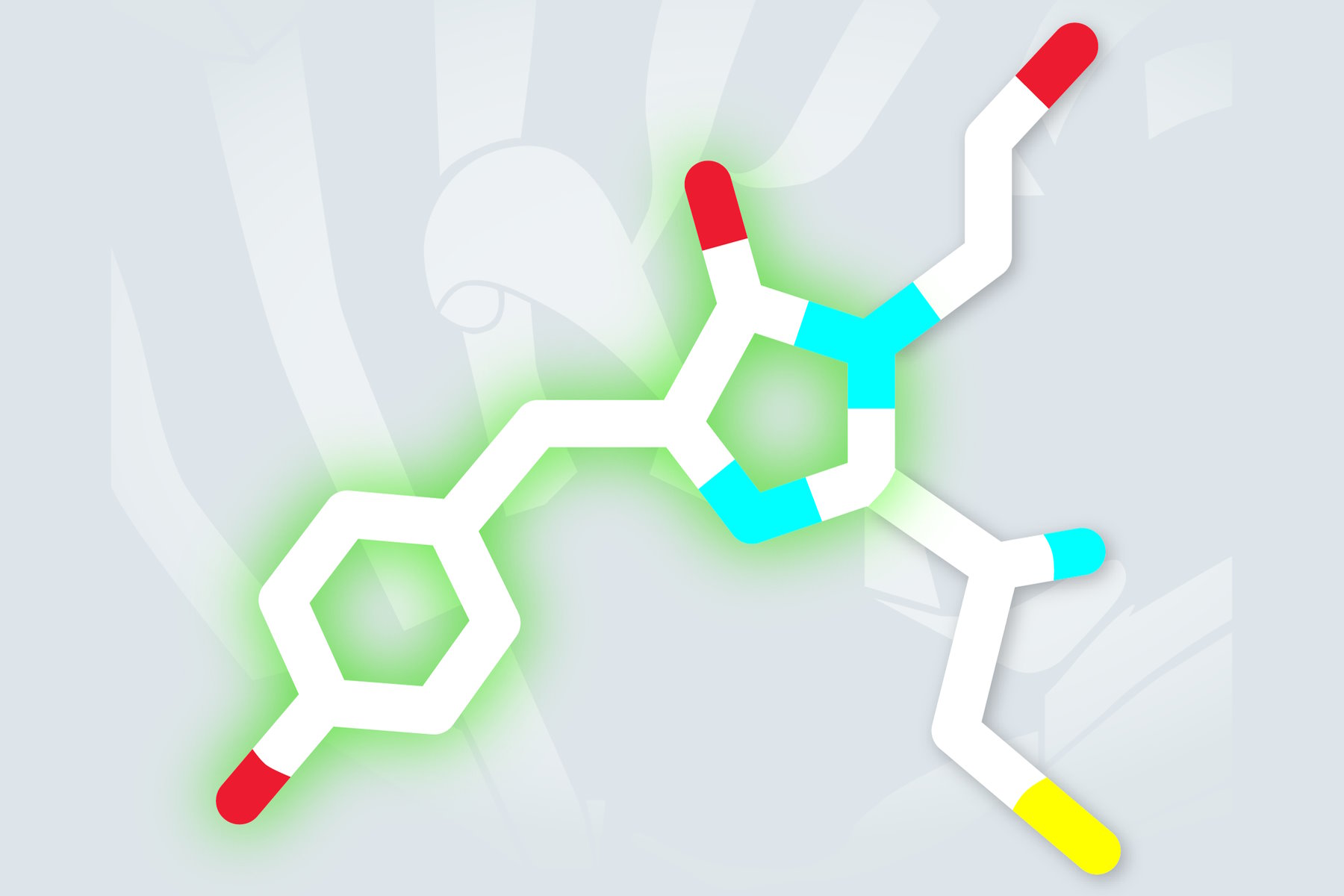
The first photoactivatable protein was a wtGFP variant and had been created by Lippincott-Schwarz and Patterson. It comprised a single point mutation (T203H), which lead to a very low absorbance in the area from 450 to 550 nm. Being photoactivated with the help of violet light, PA-GFP (photoactivatable green fluorescent protein) switches its absorption maximum from 400 to 504 nm (s. Figure 2). Therefore its fluorescence increases ca. 100-fold when excited with a wavelength of 488 nm, what is reflected in a sharp contrast between activated and non-activated proteins.
 can be activated by UV light from a non-fluorescent state to a fluorescent state.")
The underlying process during the activation of PA-GFP seems to be a light-induced decarboxylation of the glutamic acid side chain in residue 222. This loss of a carbon dioxide alters the chromophore configuration from a neutral to an anionic state [1].
Another candidate which should be mentioned here is Phamret (photoactivation-mediated resonance energy transfer), which is a special tandem dimeric fluorescent protein, photoactivated by FRET. Phamret is a fusion protein, consisting of one PA-GFP coupled covalently to its FRET partner ECFP. Exposure to 458 nm wavelength leads to emission of ECFP at 480 nm. Adjacent photoactivation of PA-GFP is performed with a 405 nm laser beam. After that, repeated exposure to 458 nm leads to FRET between the excited ECFP and PA-GFP, resulting in a green fluorescence. So the "inactive" and "active" form can be excited by the same laser wavelength (s. Figure 3). A negative characteristic could be the size of the protein which is consisting of two fluorescent proteins, possibly evoking sterical problems.
 is a tandem dimer composed of PA-GFP and its FRET partner ECFP.")
It is noteworthy that photoactivatable proteins often display a reduced brightness compared to EGFP and show a reduced photostability (s.Table 1).
Photoconvertible proteins
In contrast to photoactivatable proteins, photoconvertible proteins emit fluorescence already in their non-converted state. Therefore it is easier to define your ROI. Photoconversion was discovered in the stony open brain coral Trachyphyllia geoffroyi. Due to their emission attributes, which are shifting from green to red after photoconversion with UV light, the fluorescent protein was given the name Kaede like the leaves of Japanese maple trees [3]. In autumn they are turning their color from green to red. If Kaede is photoconverted with wavelengths between 380 and 400 nm, its emission maximum is shifting from 518 nm to 582 nm. This is displayed in a dramatically altered ratio between green and red fluorescence by the factor 2,000. This conversion is irreversible (s. Figure 4). Another limiting factor is the tetrameric nature of Kaede, what makes it challenging to use for live-cell imaging studies. Oligomerization of FPs can lead to misinterpretations concerning location and behavior of the tagged protein of interest (POI). Aggregation can even suppress the POI’s proper function completely.

The underlying process of photoconversion is again a light induced process. In this case a histidine residue inside the chromophore (His61-Tyr63-Gly64) is cleaved by irradiation and finally leads to a formation of a highly conjugated dual imidazole ring system. This process is connected to a fluorescence shift to red wavelengths [1].
A red to green convertible FP is coming from the coral Dendronephtytha sp. Its name and the red activatable attribute are mirrored in the expression Dendra [4]. The commercial development Dendra2 is the first monomeric red-to-green photoconvertible protein.
Another widely used optical highlighter is tdEosFP [5,7] and was primarily isolated from Labophyllia hemprichii, a stony coral. The tandem dimer can convert from green to red fluorescence after near-ultraviolet exposure and is gladly used for super-resolution microscopy, what is also true for the monomeric versions mEos and mEos2.
A photoconvertible protein with very similar characteristics to Keade was found in a third stony coral: Favia favus. After near-ultraviolet irradiation tetrameric KikGR changes its fluorescence from green to red, but both in a much brighter way than Kaede. Its commercial variant is called Kikume and can be photoconverted by multiphoton excitation with 750 nm wavelength. Like this it can be used in thick tissue specimens. Mutagenesis of KikGR lead to the monomeric form mKikGR. Together with mEos2 and Dendra2 it is an often used optical highlighter for super-resolution imaging.
Photoswitchable proteins
Whereas photoactivation is an irreversible conversion from a non-fluorescent state to a fluorescent state, photoswitchable proteins are able to shuttle between both conditions [6]. With the help of light pulses of different wavelengths these fluorescent proteins can be switched on and off for several hundred times without photo-bleaching. This phenomenon, to switch between fluorescent and dark states, is called photochromism and shows up already at the single molecule level of wtGFP, although to a very low extend.
One well known photoswitchable protein, which is used in super-resolution microscopy, was derived from a stony coral: Dronpa is a monomer and has one absorption maximum at 503 nm due to its anionic, deprotonated chromophore, and a minor absorption maximum at 390 nm due to its neutral, protonated chromophore (s. Figure 5). Whereas the anionic form has an emission maximum at 518 nm, the neutral form depicts a non-fluorescent state.

Furthermore there is a cis-trans isomerization connected with the protonation of the chromophore. In the neutral state of the chromophore the Tyr66 side chain is in a trans conformation (s. Figure 6). In its anionic state Tyr66 has a cis conformation. Upon irradiation with a 405 nm laser pulse Dronpa is forced into its fluorescent cis conformation. A 488 nm laser pulse then switches the Dronpa conformation to its non-fluorescent trans state. This cycle can be repeated several hundred times.


Figure 6: The Tyr66 side chain of the Dronpa chromophore undergoes cis-trans isomerization during photoswitching. While the trans-conformation is non-fluorescent, the cis-conformation emits fluorescence.
A tetrameric photoswitchable protein with emission maxima in the red region is Kindling FP (KFP1).
Latest efforts to create new optical highlighters came up with a protein which is combining both, photoconversion and photoswitching. IrisFP is a wtEosFP derivate with an on and off switchable state in its green fluorescent as well as in its red fluorescent conformation. In other words, upon irradiation with a high intensity 405 nm laser beam IrisFP is transferred from its green to a red emitting status. Green IrisFP can be switched off with the help of 488 nm laser light. Low intensity laser light at 405 nm switches it on again. On the other hand red IrisFP is switched off with 532 nm laser light and can be switched on again with a 440 nm laser. The adjacent construction of the monomeric form mIrisFP opened the door for further applications [2].

Other special fluorescent proteins
Besides fluorescent proteins which are susceptible to manipulation by light, there are additional proteins switchable by further triggers (s. Figure 8). For example, researchers who are interested in the ion status of living cells can choose GCaMP as a Ca2+ indicator. GCaMP changes its fluorescent behavior depending on its Ca2+ binding. Other fluorescent proteins such as VSFP1 are used to detect membrane potentials, whereas the yellow fluorescent protein YFP-H148Q is sensitive to Cl- ions. Furthermore there are FPs changing their fluorescent characteristics depending on the pH of their surroundings. Superecliptic pHluorin (SEP) and pHuji are two examples. With their help endo- and exocytosis events can be tracked as well as intracellular sorting events.
Fluorescent proteins which are changing their emission spectrum by time are called fluorescent timers. This behavior is not the result of pH change, ionic strength or protein concentration but happens independently from these biochemical parameters. In other words the age of the relevant protein can be estimated by its color. With these characteristics it is possible to measure time – and time depending events – inside a living cell.

The first fluorescent timer was described by the lab of Sergey A. Lukyanov in the year 2000. This DsRed mutant named FT (for fluorescent timer) alters its emission spectrum from red to green wavelengths during 18 hours. Due to the fact that FT was a tetramer Vladislav Verkhusha saw the necessity to create a monomeric one. Mutagenesis of mCherry lead to 3 more fluorescent timers with different transformation times (s. Table 1). These proteins are turning from blue to red in fast, medium or slow time scales but still too slow to measure cellular processes happening during minutes.
Nevertheless, with the help of fluorescent timers it is possible to watch the behavior of proteins in living cells, both in spatial and temporal resolution. For example differentiation and cellular polarization processes can be observed. Moreover protein transport events or viral assembly can be tracked or gene activity can be monitored. This list of applications surely could be extended and shows the potential use of Fluorescent Timers in the future.
Table 1: Selected photoactivatable, photoconvertible and photoswitchable proteins and Fluorescent Timers
Protein | Exmax(nm) | Emmax(nm) | QY | Brightness | pKa | Structure | Miscellaneous |
Photoactivatable |
|
|
|
|
|
| Condition of activation |
PA-GFP | 400 | 515 | 0.13 | 2.7 |
| Monomer | violet |
PAmCherry | 404 564 | ND 595 | ND 0.46 |
8 |
6.3 | Monomer | 350 – 400 nm |
PATagRFP | ND 562 | ND 595 | ND 0.38 | ND 25.1 | ND 5.3 | Monomer | violet |
PAmKate | ND 586 | ND 628 | ND 0.18 | ND 4.5 | ND 5.6 |
| 405 nm |
Phamret (PA-GFP + ECFP) | 458 | 475 | 0.40 | 13 |
| Monomer | violet |
Photoconvertible |
|
|
|
|
|
| Condition of conversion |
mClavGR2 | 488 566 | 504 583 | 0.77 0.53 | 14.6 17 | 8 7.3 | Monomer | 405 nm |
mMaple | 489 566 | 505 583 | 0.74 0.56 | 11.1 16.8 | 8.2 7.3 | Monomer | 380 nm |
Dendra2 | 490 | 507 | 0.50 | 22 | 6.6 | Monomer | Initial state |
PS-CFP2 | 400 490 | 468 511 | 0.2 0.23 | 8.6 10.8 |
| Monomer | violet |
Meos3.2 | 507 572 | 516 580 | 0.7 0.55 | 53 18 | 5.4 5.8 | Monomer | 405 nm |
Kaede | 508 | 518 | 0.88 | 87 | 5.6 5.6 | Tetramer | Initial state |
EosFP | 506 571 | 516 581 | 0.7 0.55 |
|
| Tetramer |
|
mEosFP | 505 569 | 516 581 | 0.64 0.62 |
|
| Monomer |
|
mEos2 | 506 | 519 | 0.84 | 47 | 5.6 | Monomer | Initial state |
kikGR (Kikume) | 507 583 | 517 593 | 0.7 0.65 | 37.6 22.8 | 7.8 5.5 | Tetramer | 365 nm |
PSmOrange | 548 636 | 565 662 | 0.51 0.28 | 58 9 | 6.2 5.6 | Monomer | blue-green |
PSmOrange2 | 546 619 | 561 651 | 0.61 0.38 | 31.1 7.2 | 6.6 5.4 | Monomer | 489 nm |
mKikGR | 505 | 515 | 0.69 | 34 | ND | Monomer | Initial state |
Photoswitchable |
|
|
|
|
|
| Condition of activation / quenching |
mTFP0.7 | 453 | 488 | 0.5 |
|
| Monomer | 405 nm / 458 nm |
PDM1-4 | 503 | 517 |
|
|
|
| 405 nm / 488 nm |
Dronpa | 503 | 518 | 0.85 | 80.8 | 5.0 | Monomer | violet / blue |
Dronpa-2 | 489 | 515 | 0.28 |
|
| Monomer | 405 nm / 488 nm |
Dronpa-3 | 489 | 515 | 0.33 |
|
| Monomer | 405 nm / 488 nm |
bsDronpa | 460 | 504 | 0.5 |
|
| Monomer | 405 nm / 488 nm |
Padron | 503 | 522 | 0.64 |
|
| Monomer | 503 nm / 405 nm |
Padron0.9 | 500 | 524 |
|
|
|
| 500 nm / 400 nm |
Mut2Q | 496 | 507 | 0.28 |
|
| Monomer | 405 nm / 478 nm |
rsFastLime (DronpaV157G) | 496 | 518 | 0.77 |
|
| Monomer | 405 nm / 488 nm |
rsKame (DronpaV157L) | 503 | 518 | 0.86 |
|
|
|
|
Dreiklang | 515 | 529 | 0.41 | 34 | 7.2 | Monomer | 365 nm / 405 nm |
mGeos-M | 503 | 514 | 0.85 | 43.9 | 4.5-5 | Monomer | 405 nm / 488 nm |
EYQ1 | 510 | 524 | 0.72 |
|
| Monomer | 405 nm / 514 nm |
KFP1 | 590 | 600 | 0.07 |
|
| Tetramer | 532 nm / 458 nm |
rsCherry | 572 | 610 | 0.02 |
|
| Monomer | 550 nm / 450 nm |
rsCherryRev | 572 | 608 | 0.005 |
|
| Monomer | 450 nm / 550 nm |
rsTagRFP | 567 | 585 | 0.11 |
|
| Monomer | 445 nm / 570 nm |
mApple | 568 | 592 | 0.49 |
|
| Monomer | 480 nm / 570 nm |
asFP595 | 572 | 595 | <0.001 |
|
| Tetramer | 569 nm / 450 nm |
Kindling FP (KFP1) | 580 | 600 | 0.07 | 4.1 | ND | Tetramer | green / 450 nm |
rseGFP | 493 | 510 | 0.36 | 16.9 | 6.5 | Monomer | 405 nm / 488 nm |
rseGFP2 | 478 | 503 | 0.3 | 18.4 | 5.8 | Monomer | 408 nm / 478 nm |
Photoconvertible/ |
|
|
|
|
|
|
|
IrisFP | 488 | 516 | 0.43 | 22 |
| Tetramer |
|
mIrisFP | 486 | 516 | 0.54 | 25 | 5.4 | Monomer | violet (dark to green) |
Fluorescent |
|
|
|
|
|
| Transition time (h) |
Slow-FT | 402 | 465 | 0.35 | 12 | 2.6 | Monomer | 9.8 |
Medium-FT | 401 | 464 | 0.41 | 18 | 2.7 | Monomer | 1.2 |
Fast-FT | 403 | 466 | 0.30 | 15 | 2.8 | Monomer | 0.25 |
mK-Go | 500 | 509 | ND | ND | 6.0 | Monomer | 10 |
Exmax: Excitation maximum, Emmax: Emission maximum, QY: Quantum Yield, Brightness is calculated by the product of Quantum Yield and molar extinction coefficient divided by 1,000
References
- Olesya V, Stepanenko O, Stepanenko V, Shcherbakova DM, Kuznetsova IM, Turoverov KK and Verkhusha VV: Modern fluorescent proteins: from chromophore formation to novel intracellular applications. BioTechniques 51 (2011) 313–327.
- Wu B, Piatkevich KD, Lionnet T, Singer RH and Verkhusha VV: Modern fluorescent proteins and imaging technologies to study gene expression, nuclear localization, and dynamics. CurrOpin Cell Biol 23:3 (2011) 310–317.
- Ando R, Hama H, Yamamoto-Hino M, Mizuno H and Miyawaki A. An optical marker based on the UV-induced green-to-red photoconversion of a fluorescent protein. Proc Natl Acad Sci USA 99:20 (2002) 12651–6.
- Gurskaya NG, Verkhusha VV, Shcheglov AS, Staroverov DB, Chepurnykh TV, Fradkov AF, Lukyanov S and Lukyanov KA. Engineering of a monomeric green-to-red photoactivatable fluorescent protein induced by blue light. Nat Biotechnol 24:4 (2006) 461–5. Epub 2006 Mar 19.
- Wiedenmann J, Ivanchenko S, Oswald F, Schmitt F, Röcker C, Salih A, Spindler KD, Nienhaus GU. EosFP, a fluorescent marker protein with UV-inducible green-to-red fluorescence conversion. Proc Natl Acad Sci USA. 101:45 (2004) 15905–10. Epub 2004 Oct 25.
- Zhou X and Lin MZ: Photoswitchable Fluorescent Proteins: Ten Years of Colorful Chemistry and Exciting Applications. Curr Opin Chem Biol. 2013 Aug; 17(4): 682–690.
- Jörg Wiedenmann, Sergey Ivanchenko, Franz Oswald, Florian Schmitt, Carlheinz Röcker, Anya Salih, Klaus-Dieter Spindler, and G. Ulrich Nienhaus: EosFP, a fluorescent marker protein with UV-inducible green-to-red fluorescence conversion
Read more about fluorescent proteins and dyes
- Fluorescent Proteins – Introduction and Photo Spectral Characteristics
- Fluorescent Dyes – An Overview
- Modern Fluorescent Proteins and their Biological Applications
- Fluorescent Proteins – From the Beginnings to the Nobel Prize

")

